




Quentin Thommen
Axes de recherche
Axes de recherche actuels
Dynamique du stress thermique en cellule unique
Collaborateurs principaux : M. Guilbert, E. Courtade ; B. Pfeuty ; F. Anquez (PhLAM)
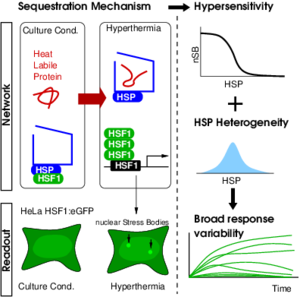
Nous étudions en particulier le rôle de la variabilité dans le destin cellulaire lors d’une perturbation de l’environnement (élévation de la température ou agression chimique) par une approche expérimentale et de modélisation. L’augmentation de la température, par exemple, engendre, au sein de la cellule, une rupture des interactions faibles, comme les liaisons hydrogène, qui stabilisent le repliement spatial des protéines : c’est la dénaturation. Les protéines dépliées s’agglomèrent, induisant une perte de l’activité biologique ce qui peut conduire à la mort cellulaire. Les protéines chaperons, qui contiennent peu de liaisons hydrogène et sont donc thermiquement stables, permettent de prévenir la dénaturation en renforçant la structure ternaire des protéines et de restaurer leurs bonnes conformations.
Même dans une population de cellule monoclonal (génome identique) on observe un continuum de réponse quant à l’activation des mécanismes intra cellulaire de protection et une mortalité partielle de la population (certaines cellules survivent au stress, d’autres non). Expérimentalement, nous suivons, par video microscopie, une population de 500 cellules sur une semaine avec une résolution temporelle de l’ordre d’une minute. Après quantification des images par une segmentation et une analyse d’image automatisée, nous obtenons le suivi de l’activation de la réponse cellulaire et du destin (mort et type de mort). Nous disposons ainsi de séries temporelles pour chaque cellule de la population, permettant d’étudier expérimentalement la variabilité. L’établissement, à partir des séries temporelles, d’un modèle de réseau de régulation guide l’étude concernant l’origine de la variabilité.
Comprendre et contrôler la variabilité cellulaire est un enjeu majeur des thérapies anti-cancer. Dans ce cadre, le stress est employé pour éradiquer les cellules cancéreuses, mais à cause de la variabilité cellulaire, il reste toujours une fraction de cellules résistantes qui re-profilèrent. Par conséquent, la capacité de finement contrôler la variabilité et la mort cellulaire, guidé par un modèle dynamique prédictif, doit permettre une optimisation des traitements. Dès lors, nos travaux s’adressent autant à la communauté de biologie et plus particulièrement de biologie systémique pour les aspects « cellule unique » et « origine de la variabilité de réponse », qu’à la communauté des praticiens pour l’aspect « nouvelles perspectives thérapeutiques ».
Optofluidique
Collaborateurs principaux : E. Courtade, J. Pesez, M. Layachi (PhLAM) ; A. Treizebré (IEMN) ; K. Braekmans (U. Gent)
Cet axe de recherche s’articule autour de deux activités expérimentales consistant à agir par la lumière de manière sélective sur des cellules humaines vivantes circulant à haut débit (∼3000 cellules par seconde), en file indienne, au centre d’un canal microfluidique (∼μm de large). Les cellules sont analysées en temps réel, une par une, à partir d’une mesure de fluorescence. Cette analyse conditionne l’activation ou pas d’un effecteur optique visant soit à (1) altérer la viabilité cellulaire (2) défléchir la trajectoire des cellules ou (3) incorporer un agent ou une molécule.
L'un des projets ,conduit dans le cadre le l’ERC Nanobubble (portée par K. Braekmans à U. Gent), vise la photoporation à haut débit dans un canal microfluidique. Des nanobulles sont optiquement générées au sein des fluides de gainage entourant les cellules dans le canal microfluidique, l’effondrement des nanobulle et l’onde de choc qui s’en suit, permet l’ouverture transitoire de nanopore dans la membrane cellulaire et donc l’incorporation intracellulaire d’un agent ou d’une molécule du milieu extracellulaire sans modification de la viabilité cellulaire. L’un des objectifs applicatifs de ce projet est l’amélioration des thérapies génétiques.
Axes de recherche en veille
Dynamique de la transcription et du gène auto-réprimé
Collaborateurs principaux : M. Lefranc et B. Pfeuty (PhLAM)
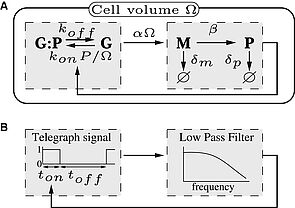
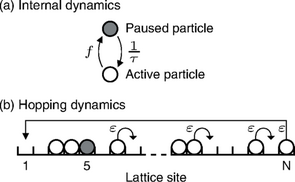
Selon le dogme central de la biologie moléculaire, l’ADN produit une molécule intermédiaire l’ARN messager (étape de transcription) servant de matrice à la production des protéines (étape de traduction) codant pour le gène considéré. La transcription est une étape complexe durant laquelle plusieurs enzymes (ARNs polymérase) se déplacent le long de la séquence de nucléotide de l’ADN en effectuant des pauses, de durée variable, le temps d’effectuer un réarrangement structurel causant, du fait de l’exclusion, des embouteillages d’enzymes. Par ailleurs, l’étape de transcription est aussi régulée en amont par des interactions entre ADN et protéines permettant ainsi la différenciation fonctionnelle des cellules et formant des circuits de régulation génétique. La régulation du gène par la protéine qu’il produit (on parle auto–régulation) est un motif fréquemment rencontré dans la nature et a été largement étudié.
En utilisant les outils de la physique statistique et de la dynamique non-linéaire (TASEP, Equation maitresse, Analyse de bifurcation) nous avons montré l’interférence entre la dynamique d’exclusion et la dynamique de pause conduisant aux embouteillages d’ARN polymérase et à la modification du taux de transcription, ainsi que l’oscillation spontanée du gène auto réprimé en présence mécanisme de dégradation nonlinéaire ou de fluctuation stochastique de la transcription.
Horloge Circadienne
Collaborateurs principaux : M. Lefranc et B. Pfeuty (PhLAM) ; F.Y. Bouget (LOMIC -UMR 7621 CNRS-UPMC)
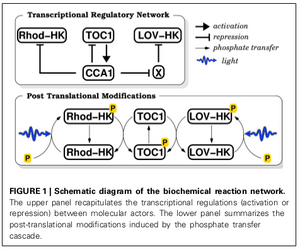
L’alternance jour/nuit induit des variations périodiques de l’environnement (lumière, tem-pérature) que la plupart des organismes vivants, dont l’homme, anticipent grâce à une horloge interne dite circadienne. Ces horloges tournent sur une période proche des 24 heures que dure le cycle lumière/obscurité. Les rouages de ces horloges sont des gènes qui interagissent afin de moduler réciproquement leur activité : par exemple, un gène « A » produit des protéines qui activent un gène « B », le gène « B » produit à son tour des protéines qui quand elles sont actives, inactivent le gène « A », et ainsi de suite. L’activité de ces gènes oscille ainsi spontanément, avec une période proche des 24 heures. Dans la cellule, on peut ainsi avoir une indication de l’heure qu’il est en mesurant la concentration de ces protéines et donc leur activité.
À partir de mesures expérimentales conduitent par l’équipe de F.Y. Bouget de l’Observatoire océanologique de Banyuls (CNRS/UPMC) sur l’horloge de l’algue verte unicellulaire Ostreococcus tauri. Nous avons étudié au travers d’un modèle mathématique, les propriétés de synchronisation robuste de l’horloge sur le cycle jour/nuit.
Axes de recherche développés durant la thèse et le postdoctorat 2001--2006
Dynamique quantique dans les potentiels lumineux
Sujet de recherche initié dans le cadre de mon doctorat en collaboration avec Jean-Claude Garreau et Véronique Zehnlé (PhLAM).
Les techniques de refroidissement et de piégeage d'atomes par lasers permettent d'obtenir des gaz atomiques de très faible densité à des températures de l'ordre de quelques micro-Kelvin. L'utilisation de champs lasers permet d'appliquer aux atomes des potentiels mécaniques variés (on parle alors de ``potentiels lumineux''), comme par exemple un réseau incliné, , somme d'un terme spatialement périodique et d'un terme linéaire: , potentiel intéressant de par sa double symétrie de translation espace--énergie. La dynamique quantique d'un atome dans un réseau incliné est connue depuis les travaux de Zener en 1928. Elle n'en demeure pas moins d'actualité: il s'agit d'une oscillation sur plusieurs puits de potentiel nommée: les oscillations de Bloch. Cette dynamique purement quantique traduit l'existence d'une fréquence naturelle dans le réseau incliné.
- Dynamique quantique dans un réseau incliné
Par une étude analytique, nous avons montre que la modulation harmonique des paramètres du réseau incliné induit un phénomène de résonance entre la fréquence de modulation et la fréquence naturelle induisant un transport cohérent de la particule dans le réseau incliné: translation et/ou diffusion. Ce transport est pilote par les cohérences quantiques initiales de l'atome tandis de la vitesse de déplacement est proportionnelle à l'amplitude de modulation.
Dans le cas d'un réseau bidimensionnel, la modification lente des amplitudes de modulation dans les deux directions de l'espace permet d'asservir temporellement la vitesse de la particule et de générer une grande variété de trajectoires de l'atome dans le plan, réalisant ainsi un moteur quantique. Similairement la modification spatiale de l'amplitude de modulation permet de concentrer un nuage atomique ou d'induire une rotation dans le plan de la fonction d'onde atomique.
- Dynamique d'un condensat de Bose Einstein dans un réseau incliné
La condensation de Bose Einstein est l'accumulation macroscopique de particules dans l'état de plus basse énergie d'un puits de potentiel. Le gaz d'atomes condensés peut alors être considéré comme une particule unique. L'évolution dynamique d'un {condensat de Bose Einstein} dans le réseau incliné sans modulation temporelle est beaucoup plus riche et variée que l'évolution d'un atome, de par la non--linéarité introduite par la description en champ moyen des interactions entre particules.
Nous avons montré que la dynamique quantique du condensat de Bose Einstein est décrite par une trajectoire classique dans cet espace de phase ad hoc et correspond à celle d'un système hamiltonien quasi-intégrable, incluant une évolution temporelle chaotique (au sens classique) du condensat. L'analogie formelle démontrée entre la dynamique quantique du condensat dans un réseau incliné et la dynamique d'une chaîne d'oscillateurs non-linéaires classiques couplés est un résultat nouveau et remarquable,
- Reconstruction de la fonction d'onde {\it via} les oscillations de Bloch
Enfin, nous avons établi une méthode, utilisant les oscillations de Bloch et permettant la détermination complète de la fonction d'onde atomique. Si le principe d'indétermination de Heisenberg nous interdit de définir conjointement position et vitesse de manière exacte il n'y a, en effet, aucune limite à la connaissance de la fonction d'onde. L'expression de la fonction d'onde dans l'espace réel joue un rôle prépondérant dans notre compréhension de la mécanique quantique, et la mise en oeuvre d'une technique expérimentale permettant de la reconstruire est d'un grand intérê: si la distribution de probabilité spatiale est souvent facilement mesurable, la mesure de la phase nécessite généralement l'emploi d'une méthode annexe et souvent incompatible, car la mesure de la position d'un système quantique efface généralement l'information sur la phase. Notre méthode, qui est basée sur la mesure d'une classe de vitesse, est aussi applicable au cas d'un condensat de Bose Einstein et permet de déterminer, outre la fonction d'onde du condensat, le paramètre non-linéaire.
Matériaux gauchers induits par cohérences quantiques
Sujet de recherche développé dans le cadre de mon post--doctorat en collaboration avec Paul~Mandel de l'Université libre de Bruxelles --- ce sujet n'est plus actif pour le moment
En 1964, Veselago étudie théoriquement la propagation des ondes électromagnétiques dans des milieux matériels ayant une permittivité et une perméabilité réelles et négatives. L'indice de réflexion est alors réel et le milieu propage les ondes électromagnétiques. Le fait marquant est qu'en étudiant les conditions de passage des champs, Veselago montre qu'il faut attribuer à ces matériaux un indice de réflexion négatif. De plus, dans ces matériaux, le trièdre formé par les vecteurs champ électrique, excitation magnétique et le vecteur d'onde est indirect, c'est-à-dire qu'il suit la règle de la main gauche. De tels milieux ont alors été nommés ``matériaux gauchers''.
En 2000, Pendry démontre que les matériaux gauchers permettent de focaliser les ondes électromagnétiques en deçà de la longueur d'onde du rayonnement en amplifiant les ondes évanescentes. Les matériaux gauchers ouvrent ainsi la voie vers la réalisation de "lentilles parfaites". Depuis, motivées par les applications technologiques, l'étude et la réalisation de matériaux gauchers sont l'objet d'efforts de recherche considérables. L'intérêt des matériaux gauchers est double, à la fois théorique et pratique. Expérimentalement, le premier matériau gaucher a été obtenu dans le domaine micro-ondes par l'emploi d'un réseau de résonateurs électromagnétiques métalliques.
Notre démarche est d'utiliser des transitions atomiques comme des résonateurs électromagnétiques afin de modifier les constantes électromagnétiques dans un domaine spectral proche des fréquences de transition: une transition dipolaire électrique joue le rôle du résonateur électrique et une transition dipolaire magnétique joue le rôle du résonateur magnétique.
À partir d'un schéma général impliquant quatre niveaux atomiques, nous avons présenté la première mise en évidence théorique de l'existence de domaines de paramètre gauchers dans un gaz atomique. L'absorption a été réduite en jouant sur les interférences quantiques entre amplitudes de transition. Par ailleurs en utilisant les propriétés spectroscopiques de l'ion erbium placé dans une matrice cristalline, nous avons présenté un schéma différent permettant la première mise en évidence théorique de propriétés gauchères d'un cristal dues aux cohérences quantiques. Il est remarquable que le domaine gaucher soit centré autour de la longueur d'onde utilisée pour les transmissions de télécommunications (1.54µm).